MENDEL
Y SU OBRA
Autor:
Ing. Agr. Carlos González
Johan Gregor Mendel
(1822-1884) nació en Kuclanchen, pequeña aldea austríaca. Hijo de una familia
campesina, ayudaba a su padre, fruticultor, en trabajos de granja y huerta. Al
no poder completar sus estudios en la Universidad de Viena, debió dedicarse al
seminario en el monasterio de Brünn (Hoy Brno), en la Rep. Checa. En él cursó
estudios teológicos, ordenándose sacerdote a los 24 años. Llegó a ser prelado
del mismo monasterio. Allí se dedicaba a cultivar flores en el jardín, y para
aumentar sus conocimientos realizó cursos en la Universidad Local.
|
Así comenzó sus
investigaciones sobre leguminosas, descubriendo las leyes que dieron origen a
la Genética. Trabajando por 8 años, dónde efectuó recuentos y conclusiones
estadísticas, que fueron presentados en 1865 en la Sociedad de Historia
Natural de Brünn, frente a un auditorio que no le dio importancia. Al año
siguiente publicó en las Actas de dicha sociedad su monografía de "Experimentos
sobre la hibridación de las plantas", quedando ignorada durante más
de treinta años. Permaneció en el olvido hasta 1900, que fue redescubierta
por tres botánicos en forma independientes (El alemán Correns, el holandés De
Vries y el austríaco Von Tcherman). |
|

Es conveniente
notar que Mendel, trabajando con material semejante al utilizado por otros
investigadores anteriormente, obtiene resultado a causas de varias razones:
a)El material
escogido: Eligió las plantas leguminosas, y entre ellas el chícharo de
jardín o arvejilla de olor (Pisum sativum), que presenta una serie de
ventajas:
- Fácil de cultivar en macetas y canteros.
- Ciclo vegetativo corto.
- Autofecundación natural y fecundación cruzada
difícil (El estilo está escondido por los pétalos).
- Producción de gran cantidad de semillas por planta,
lo que le permite obtener gran cantidad de descendientes, especial para el
estudio estadístico.
- De fácil obtención: el conseguía en el comercio.
- Gran cantidad de variedades, con caracteres
simples, variados y contrastantes.
El estudió solo siete pares,
ubicados en distintos cromosomas (fig. 4):
|
|
|

b)Método
Mendeliano:
La razón principal
de sus éxitos:
- Tomó caracteres simples, contratantes y visibles
y los analizó independientemente, evitando de esta manera confundir los
resultados.
- Para cada caso estudió varias generaciones sin
interrupción.
- Resolvía una cuestión por vez para luego encarar
cuestiones más complejas (estudió primero los monohíbridos, luego
dihíbridos y a posterior trihíbridos, etc)
- Contó la prole y después analizó los resultados
matemáticamente (estadísticamente). Lo más importante.
- Establecía hipótesis, verificando si coincidían
con los hechos, sin limitarse a una coincidencia cualitativa.(Estableció
un método lógico).
- LEYES:
Mendel estableció
tres principios (denominadas leyes) fundamentales basados en los resultados
obtenidos por sus experimentos:
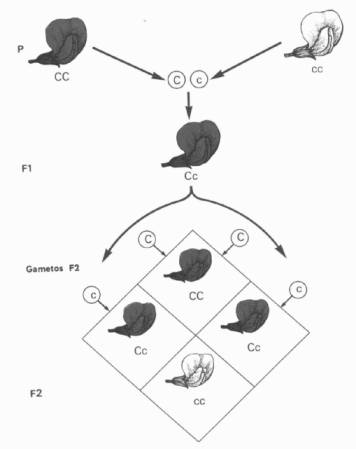
PRINCIPIO DE LA UNIFORMIDAD (Primera ley)
Tomando un par de caracteres
simples, contrastantes y bien visibles, como el caso de las plantas altas y
bajas, Mendel las cultivó separadamente en canteros, mediante autofecundación,
durante varias generaciones, dejando sólo aquellas que producían individuos con
las características paternas(color de flor en este caso), los consideró puros
para el carácter. Una vez obtenidas las plantas puras para flores rojas y
blancas respectivamente, verificó el resultado del cruzamiento de esos dos
tipos de plantas, con los siguientes resultados:
a)Las semillas
obtenidas, producto del cruzamiento, dieron plantas hijas que en su totalidad
eran rojas. Es decir un 100 % de la descendencia era de pie roja (era
como si el carácter flor blanca no existiera (Primera Generación o Filial 1 o
F1).
|
Filial
1 y filial 2 |
PRINCIPIO
DE SEGREGACION (Segunda
ley o de Separación Monofactorial o de separación de factores):
b) Si cruzaba
entre sí a los individuos de flor roja de esta primera generación (o los dejaba
que se autofecunden) resultaban semillas que darían plantas de flor roja y
blanca en una proporción 3:1, o sea un 75 % de flor roja y un 25 %
de flor blanca (Segunda Generación o
Filial 2 o F2).
c) Si se cruzaba
entre sí, a las plantas de flor blanca obtenidas en la segunda Filial, o se autofecundaban,
todos los descendientes eran siempre blancas, por lo tanto puros para ese
carácter. Exactamente igual a uno de los progenitores.
d) Si se
autofecundaban las plantas de la F2 de flore rojas o si se cruzaban
entre sí resultaba:
- La tercera parte
de ellos (1 de cada 3) daban individuos de flores rojas únicamente, durante las
generaciones subsiguientes, es decir que se comportaban como puros para ese
carácter, igual al otro de los progenitores.
- Las otras dos
terceras partes, que eran de flores rojas, daban una descendencia de flores
rojas y blancas en una proporción 1:3 (75 % y 25 %), es decir se comportaban
como los impuros (igual a las plantas de la primera generación. En
cuanto a la pureza de los individuos para dicho carácter, las generaciones
siguientes era compuestas por una relación 1(flores rojas puro) : 2 (flores
rojas impuro):1 (flores blancas puro).
Mendel repitió
esta experiencia varias veces y contó a miles de plantas, también lo hizo con
otras características (como la forma de la semilla, el color, tipo de vainas,
etc.)obteniendo siempre las mismas proporciones, es decir una primera
generación uniforme para uno de los caracteres paternos y en la segunda una
proporción de 3 a 1 (tres iguales a los de la primera generación y uno
semejante al padre que había desaparecido dicho carácter).Y 1:2:1 en cuanto a
pureza.
Estableció
entonces los siguientes hechos como constantes y generales:
1) Los productos
de la primera generación (F1) presentaban las características de uno
de sus progenitores, a este factor lo denominó dominante, mientras el otro parece haber desaparecido.
2) En los
productos de la segunda generación (F2), reaparece el carácter
oculto en un individuo de cada cuatro(25 %). Dedujo que factor que determinaba este
carácter estaba en la primera filial (F1)pero oculto, por eso lo
llamó recesivo. Los descendientes de la F2 fueron siempre
iguales, por lo que dedujo que eran puros.
3) Si dejaba
autofecundar los productos de la F2 del factor dominante (las tres
cuartas partes): roducen descendientes de las mismas características, por lo
tanto son "puros", mientras que dos terceras partes se comportaban
como la F2 (producían 3:1), por lo tanto eran "impuros".
4) A los
dominantes y recesivos puros, se los llamó homocigotas, porque sus cigotos son puros y sólo producen gametos
con el carácter de uno de sus progenitores. A dominantes impuros, se los
denominó heterocigotas, por tener los caracteres de ambos progenitores.
Distribución de
las gametos:
Al formular la
hipótesis, Mendel, consideró que las células sexuales de las plantas, óvulo y
polen eran agentes que transportaban los factores hereditarios que determinaban
la aparición de las características de la descendencia. Puesto que cuando un
individuo puro para un carácter dominante o recesivo, producirá gametos
masculinos y femeninos estos llevarán el mismo factor determinante. En el caso
de las arvejas de semilla amarilla (YY) sus gametos (Y)serán también de semilla
amarilla(tanto masculinos como femeninos).Otro tanto sucede con las plantas de
semilla verde (yy). Al cruzar estos individuos puros, entre sí(las dos líneas),
cada uno de los padres aportará ya sea en el polen o en el óvulo, el carácter
que le corresponde (amarillo-Y o verde-y-). Como la fecundación consiste en la
fusión de polen y óvulo para formar la célula huevo, el nuevo embrión contendrá
los dos factores(individuo impuro o híbrido: Yy), para formar la nueva planta y
todas sus células lo contendrán. La dominancia de uno de ellos (amarilla:Y),
hace que se manifieste esto y el otro se mantenga oculto(recesivo: y). Si bien
Mendel desconocía los fenómenos de la meiosis, supuso que al producirse las
nuevas gametas, estos factores nunca entraban junto en la célula reproductora.
Esta separación o segregación de los factores hereditarios es el principio
fundamental de la hipótesis mendeliana.
Según esta idea,
el híbrido producía granos de polen de dos clases, la mitad con el factor dominante (Y) y la otra con el recesivo (y). De la misma manera se produciría dos clases de
óvulos. Al realizarse la fecundación, estos volverían a reunirse. De tal manera
que si autofecundamos estos tendrías las siguientes combinaciones:
- Que un óvulo con
factor dominante (Y) se cruce con polen de factor dominante (Y), lo que daría un
individuo dominante puro (YY).
- Que un óvulo con
factor dominante (Y) se cruce con polen de factor recesivo (Y), lo que daría un
individuo dominante impuro (Yy).
- Que un óvulo con
factor recesivo (y) se cruce con polen de factor dominante (Y), lo que daría un
individuo dominante impuro(Yy).
- Que un óvulo con
factor recesivo (y) se cruce con polen de factor recesivo (y), lo que daría un
individuo recesivo puro (yy).
Por lo que
tendrías tres plantas que manifiestas el factor dominante (una pura y dos
híbridas) y una el factor recesivo que es pura.
Los resultados
experimentales obtenidos por Mendel corroboran esta aseveración, que explica
perfectamente la proporción 3:1 de las características externas, y la relación
1:2:1 con los individuos impuros.
Como es necesario
reconocer que la fusión de las gametas se hace completamente al azar,
sin que exista ninguna afinidad que haga que un gameto de cierta clase se una
otro de clase determinada, todas las combinaciones son igualmente posibles, por
lo tanto de igual número. Esta combinación libre de los factores es el
fundamento de la segunda ley de Mendel.
El concepto de
carácter unitario, como algo separable e independiente, es el logro básico de
Mendel, y según sus observaciones, para cualquier carácter dado se podría
encontrar otro contrastante y con él formar un par de caracteres contrastantes
(o alternativos). Actualmente a este par se lo denomina par de alelos. Un miembro puede tener para un carácter un par de
alelos, es decir un gen con dos alternativas diferentes en cada uno (ejemplo el
carácter de semilla tegumento de color verde es una alternativa y su par
alelomorfo es el tegumento de color blanco).
Al carácter
externo del individuo que se somete a estudio, se lo denomina carácter fenotípico(o fenotipo, que por extensión se lo denomina al grupo
de individuos con carácter externos comunes).En el ejemplo la semilla amarilla
es el fenotipo, aunque sabemos que hay individuos puros (YY) o impuros (Yy).A
esta característica que no se manifiesta pero que representa la constitución
genética del carácter o individuo se lo denomina carácter genotípico (o genotipo). En nuestro caso de la F2 los
fenotipos serían 1 planta de semilla amarilla pura (YY), dos de semilla
amarilla impuras (Yy) y uno de semilla amarilla pura (yy). La relación
existentes entre los individuos de una filial, denominamos frecuencia
fenotípica, si se refiere a los fenotipos (3:1, en el ejemplo) o frecuencia
genotípica, cuando nos referimos a los genotipos (1:2:1).
Para esquematizar,
lo organizamos en un esquema dónde a la generación paterna inicial lo
representamos como P (padres), a la primera generación como F1
(filial uno), a la segunda generación F2 (filial 2) y las
gametas G. En otro ejemplo sería:
P DD
(pie alto) x dd (pie bajo)
G D d
F1 Dd
F1 x F1 Dd x Dd
F2 DD Dd Dd dd
Frecuencia
genotípica: 1 : 2 : 1
Frecuencia
fenotípica: 3 : 1
Para simbolizar los genotipos se pueden
adoptar letras, optando por las letras mayúsculas para los
dominantes y las minúsculas para los recesivos (DD,
Dd, dd)tal como lo hizo Mendel. También se pueden utilizar letras con
exponentes o subíndices, muy usado en los alelos múltiples (ej: C+, cch, ch, ca, en
el pelaje de los conejos o R1, R2, r1, r2
del color de grano del maíz), como utilizó Morgan. También es muy usada la
forma fraccionaria (B/b, b/b, B/B).
PRINCIPIO DE DISTRIBUCION
INDEPENDIENTE (tercera ley de trasmisión o segregación independiente o de
independencia de los caracteres)
Mientras el principio de segregación se
aplica al comportamiento de un sólo gen, este describe el comportamiento
simultáneo de dos o más pares localizados en distintos cromosomas.
Si cruzamos dos individuos de raza pura en el
cual seleccionamos dos pares de caracteres (ubicados en dos loci) obtenemos
descendencia híbrida para los dos caracteres (dihibridos). Haciendo un análisis semejante al caso anterior
obtenemos:
a)Una generación F1 homogénea y
doblemente heterocigota.
b)Una segregación gamética, en la cual aparecen
las cuatro combinaciones posibles, cada una conteniendo un alelo de cada par de
genes, siendo la frecuencia de cada genotipo igual a 1/4.
|
|
c)Una segregación cigótica, con nueve
genotipos diferentes, siendo cuatro homocigotas dobles con frecuencia 1/16 cada
uno, cuatro heterocigotas simples con frecuencia 2/16 cada uno y finalmente un
doble heterocigota con frecuencia igual a 4/16.
|
|
Cuando Mendel interpretó el mecanismo que regía
la herencia de un par de caracteres contrapuestos o alelomorfos, continuó con
el fenómeno hereditario en el caso de individuos cruzados que difiriesen en dos
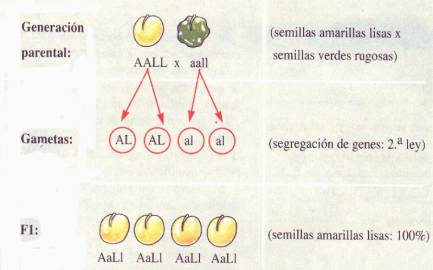
pares de caracteres. Cruzó plantas de arveja de semilla lisa y amarilla con
plantas de semilla rugosa y verde. Ambas había sido seleccionadas previamente
por ser puras por autofecundaciones sucesivas. El carácter liso (L) era
dominante sobre el rugoso (l) y el color amarillo (A) sobre el verde (a).Dicho
cruzamiento proporcionaba una F1 constituida por plantas que de
semilla lisa amarilla (L_A_), que corroboraba la dominancia de estos caracteres
de la misma manera que cuando actuaban aisladamente. Realizando cruzamientos o
autofecundaciones entre los individuos de la F1, obtenía en la F2
los tipos primitivos, es decir con los mismos caracteres de los padres, como en
la segunda ley; pero además aparecían nuevas combinaciones, no existentes en
generaciones precedentes. Las nuevas combinaciones aparecidas, en este caso,
eran plantas con semilla lisa y verde y semillas rugosa y amarilla. Haciendo el
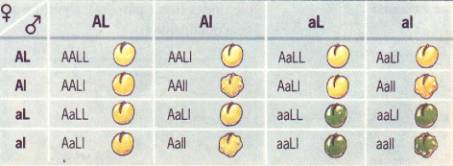
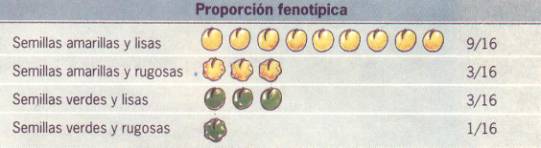
recuento de individuos en la F2, Mendel encontró que la relación
estadística de fenotipos encontrados era de 9 lisos amarillos (L_A_), 3 lisos
verdes (L_aa), 3 rugosos amarillos (llA_) y 1 rugoso verde (llaa), o sea 9:3:3:1.
|
|
En el mismo cruzamiento analizando únicamente
un par alelomorfico. Por ejemplo: Liso y rugoso (L/l)-, verificó que en la F2,
dentro de la relación 9:3:3:1 había 12 lisos y 4 rugosos, o sea 3:1 cumpliendo
con la segunda ley (en estos 1 liso es puro y dos heterocigotas). Analizando el
par amarillo y verde (A/a) llegó a la misma conclusión, también se adaptaba a
la segunda ley, como si el otro par no existiera, lo que demostró que cada par
se comporta independientemente de los otros.
|
|
|
|

Las nuevas combinaciones aparecían debido a
que en la formación de las gametas se separan los genes pertenecientes a un
mismo de caracteres alelomorficos, de manera que un gameto contiene solamente
uno de estos genes de cada par alelomorfico; pero además en este caso la
asociación en los gametos del gene correspondiente a un par con el otro par se
efectúa al azar.
En
resumen: Caracteres:
L=
liso l= rugoso A= amarillo a= verde
P LLAA x
llaa
(liso
y amarillo) (rugoso y verde)
G (LA)
(la)
F1
LlAa
(liso
y amarillo)
F1
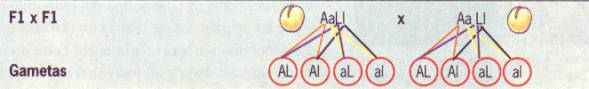
x F1 LlAa x LlAa
G
(LA)(La)(lA)(la) (LA)(La)(lA)(la)
|
F2 |
9 L_A_ |
3 L_aa |
3 llA |
1 llaa |
|
|
(liso-amarillo) |
(liso-verde) |
(rugoso-amarillo) |
(rugoso-verde) |
|
|
1 LLAA |
1 Llaa |
1 llAA |
1 llaa |
|
|
2 LlAA |
2 Llaa |
2 llAa |
|
|
|
2 LLAa |
|
|
|
|
|
4 LlAa |
|
|
|
(relación
fenotipica) 9:3:3:1
(relación
genotípica) 4:2:2:2:2:1:1:1:1
Homocigotas
doble(4), uno de cada clase(1/16)
Heterocigotas
simples(8), dos de cada clase(2/16)
Heterocigotas
doble (4), cuatro de cada clase (4/16)
De
esta manera la relación fenotípica de un dihíbrido(dos pares de factores
independientes es de 9:3:3:1, y la relación genotípica es 1:2:2:1:4:1:2:2:1
TRIHIBRIDOS Y OTROS POLIHIBRIDOS
Cuando se cruzan
entre sí individuos homocigotas que difieren en tres pares de factores, el
híbrido resultante se llama trihíbrido.En general, se denominan polihíbridos a
los heterocigotas un numero n de pares alelomóficos.
Lo señalado para
la herencia de un dihíbrido puede ser aplicados a 3 o más pares de caracteres,
pues la ley de pureza de las gametas, también se aplica.
Supongamos el
ejemplo de un trihíbrido. En conejo se observa que el pelo corto (C) es
dominante sobre el largo (c), el de color negro (N) domina al albino (n) y la
grasa blanca (B) es do-minante sobre la grasa amarilla (b).
P CCNNBB x ccnnbb
(P.corto,negro
y G.blanca) (P.largo,albino y G.amarilla)
G CNB
cnb
F1
CcNnBb
(Pelo
corto, negro y Grasa blanca)
G CNB
CNb CnB cNB cnB cNb Cnb cnb
F1 x F1 CcNnBb x CcNnBb
F2
|
27 |
9 |
9 |
9 |
3 |
3 |
3 |
1 |
|
C_N_B_ |
C_N_bb |
C_nnB_ |
ccN_B_ |
C_nnbb |
ccN_bb |
ccnnB_ |
ccnnbb |
|
P.cort |
P.cort |
P.corto |
P.largo |
P.corto |
P.largo |
P.largo |
P.largo |
|
P.negr |
P.negr |
P.albin |
P.negro |
P.albin |
P.negor |
P.albin |
P.albin |
|
G.blan |
G.amar |
G.blanc |
G.blanc |
G.amar |
G.amar |
G.blanc |
G.amar. |
(Relación
fenotípica 27:9:9:9:3:3:3:1)
|
1CCNNBB |
1CCNNbb |
1CCnnBB |
1ccNNBB |
1CCnnbb |
1ccNNbb |
1ccnnBB |
1ccnnbb |
|
2CCNNBb |
2CCNnbb |
2CcnnBB |
2ccNnBB |
2Ccnnbb |
2ccNnbb |
2ccnnBb |
|
|
2CCNnBB |
2CcNNbb |
2CCnnBb |
2ccNNBb |
|
|
|
|
|
2CcNNBB |
4CcNnbb |
4CcnnBb |
4ccNnBb |
|
|
|
|
|
4CCNnBb |
|
|
|
|
|
|
|
|
4CcNnBB |
|
|
|
|
|
|
|
|
8CcNnBb |
|
|
|
|
|
|
|
Relación
genotípica
8:4:4:4:4:4:4:2:2:2:2:2:2:2:2:2:2:2:2:1:1:1:1:1:1:1:1
que
puede abreviarse en la práctica de la siguiente manera:
(1)8:(6)4:(12)2:(8)1