Envoltura
nuclear
La envoltura nuclear consta de dos membranas que separan el contenido nuclear del citoplasma circundante. Las dos membranas de la envoltura se interrumpen en algunos puntos formando poros nucleares, de tal forma que el interior del núcleo se comunica con el citoplasma celular. Los poros nucleares presentan canales acuosos que permiten el paso de sustancias del interior de núcleo hacia el citoplasma y viceversa, pero el proceso es altamente selectivo, permitiendo solamente el paso a moléculas específicas.
El poro nuclear en realidad es una estructura altamente elaborada, denominada complejo del poro nuclear, compuesta de más de 100 proteínas diferentes, ordenadas con una simetría octogonal. Las moléculas pequeñas (5 kDa o menos) difunden en forma prácticamente libre, pero las proteínas de gran tamaño necesitan contar con un señal de localización nuclear, que generalmente consiste en una corta secuencia de aminoácidos (de 4 a 8). El proceso de entrada de una proteína destinada al núcleo necesita que otra proteína citosólica ("receptor nuclear de importación") llamada nucloporina se una a la señal de localización nuclear y requiere además de la energía que proporciona la hidrólisis de una molécula de trifosfato de guanidina (GTP). Esto provoca la dilatación del poro y permite el pasaje de la proteína. La salida de las subunidades ribosómicas fabricadas en el nucleolo y el ARNt también dependerían de un sistema de transporte activo mediado por señales de exportación nuclear.
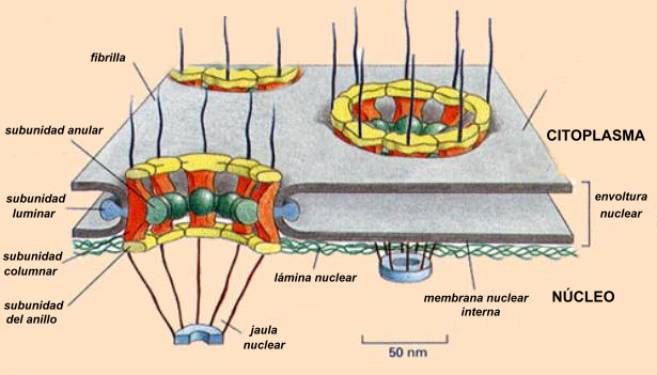
Unida al interior de la membrana nuclear se
encuentra una capa de proteínas específicas que al parecer funcionan como
esqueleto del núcleo (lámina nuclear) y que tiene un papel importante en
la desorganización y reorganización de las membranas nucleares al comienzo y al
fin de la división celular, respectivamente. La lámina nuclear es un enrejado
de subunidades proteicas, del tipo de los filamentos intermedios que han sido
vistos al tratar el citoesqueleto (proteínas fibrilares) y que, como todas las
proteínas nucleares lleva una “señal de transporte nuclear” que los dirige
desde el citosol, donde son sintetizados, hacia el núcleo.
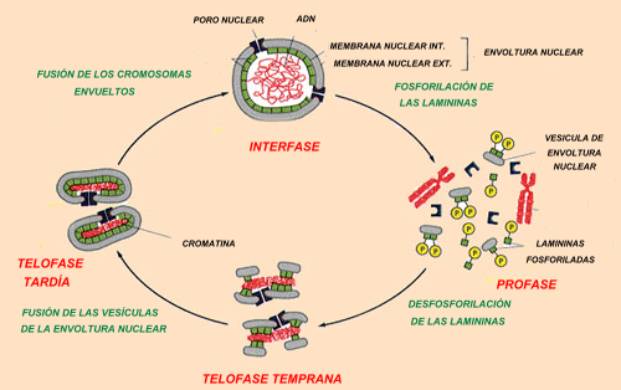
Cuando el núcleo se desensambla durante la mitosis, la lámina nuclear se depolimeriza por fosforilación. Al mismo tiempo la membrana nuclear se desarma en vesículas membranosas, que van adheridas a las proteínas fosforiladas (fracciones de la lámina nuclear). En la telofase temprana (una de las fases finales de la división celular) se produce la defosforilación de las proteínas y las vesículas se repolarizan alrededor de cada cromosoma. En la telofase tardía las vesículas se reúnen y reconstituyen la envoltura nuclear de cada célula hija, que activamente reimporta a las proteínas que llevan la señal de transporte nuclear y recompone los poros nucleares.