Los Retrovirus
![]()
Ningún otro grupo de virus ha recibido tanta atención en los últimos años como los retrovirus (RV). Los RV han mostrado peculiaridades biológicas que les hacen muy diferentes de los otros grupos de virus y que han dado lugar a la reinterpretación de algunos dogmas de la biología en vigor hasta hace bien poco. La propiedad más característica de este grupo de virus es la retrotranscripción, descubierta en 1970 por Baltimore. Este proceso consiste en la síntesis de una molécula de ADN utilizando como patrón un ARN, y es una actividad enzimática que no existe en las células normalmente. La observación de este fenómeno obligó a modificar el dogma central de la biología molecular acerca del sentido del flujo de la información genética ADN- ARN- proteína. También, en el campo de la microbiología, los postulados de Pasteur y los conceptos clásicos sobre las relaciones huésped- microorganismo se han visto limitados en su aplicación a estos agentes. Por ejemplo, el concepto de microorganismo como agente externo se ha visto replanteado por la íntima relación de estos agentes con el huésped.
Los RV son capaces de incorporar su genoma a los cromosomas de organismos superiores y de inducir cambios en el de estos. Una de las consecuencias de este fenómeno es la enorme capacidad de los RV para modificar los genes del huésped, por lo que los RV representan una importante fuerza en la evolución de los vertebrados. Su relación con el huésped es tan estrecha que los RV pueden estar presentes en circunstancias normales, por ejemplo en la placenta, y en ocasiones desempeñan papeles fisiológicos, como es el caso del desarrollo y control del sistema inmune. Quizá por todo esto, ningún otro agente es tan atractivo desde el punto de vista teórico como los RV para explicar la autoinmunidad, o lo que es lo mismo, la confusión entre lo propio y lo ajeno.
Existe, por último, otro motivo para que el estudio de los RV, que permanecía en el punto de mira de unos pocos investigadores, se haya extendido a todos los ámbitos de la medicina y la sociedad. Este es el impacto de una enfermedad devastadora, el SIDA, que ha generalizado el interés por los RV y nos ha mostrado de forma incuestionable la capacidad de estos agentes para alterar el sistema inmune.
Concepto y clasificación
Desde el punto de vista de su organización genética, los RV forman parte de los retroelementos, que incluyen varios tipos de estructuras similares. La característica común a todos ellos es la posibilidad (al menos en su origen evolutivo) de introducirse <<marcha atrás>> en el genoma de otros organismos, o lo que es lo mismo, de integrarse en el ADN genómico, tras la retrotranscripción de su ARN. La presencia de estos elementos en el genoma humano normal es tan importante que solamente un subgrupo denominado L1 contiene unas 100.000 copias, lo que supone el 5% de todo el material genético humano. Por ello, no es infrecuente encontrarlos de forma casual durante los esfuerzos por aislar genes convencionales y de esta forma han sido conocidos muchos de ellos. Dependiendo de la capacidad de sus genes de dar lugar o no a productos, existen retroelementos completamente inactivos y otros capaces de transcribirse y sintetizar proteínas. Su estructura permite que puedan copiarse e insertarse en otro punto del genoma por lo que se denominan retrotransposones, o elementos móviles en el genoma.
Un RV es un retroelemento con capacidad para ensamblar sus productos proteicas formando partículas virales. Ocasionalmente, pueden ser viables fuera del organismo y potencialmente infecciosos o por el contrario ser exclusivamente endógenos, procedentes del ADN del huésped. La viabilidad fuera de las células es una característica exclusiva de los RV determinada por la presencia de una proteína de cubierta (env) que les proporciona capacidad para unirse a sus receptores celulares, generalmente glucoproteínas de membrana. Por ello, la ausencia o defectos en el gen env les incapacita para tener vida fuera de la célula y capacidad infecciosa. Un ejemplo del ciclo completo se muestra en la figura 1 y defectos en alguno de los procesos (integración, retrotranscripción, etc.) explican las posibles diferencias entre los múltiples grupos de RV.
Un RV completo consta de dos copias iguales de su ARN genómico (son virus ARN) que contiene tres genes fundamentales: gag, pol y env. Estos codifican 3 grupos de proteínas, gag, las que forman el core o nucleocápside proteica, pal, las que realizan las actividades retrotranscriptasa e integrasa y env, las que forman la cubierta.
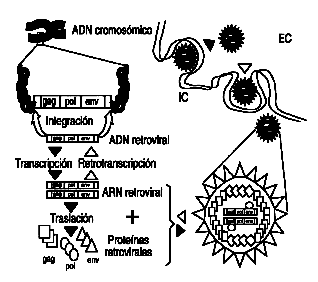
Figura 1. Ciclo celular de los retrovirus (RV). El ciclo completo de un RV infeccioso (flechas blancas) comienza tras su entrada en la célula mediante la interacción de la cubierta (env) con receptores de membrana. Desde el espacio extracelular (EC), penetra en la célula (espacio intracelular, IC) donde, por la acción de proteasas, sus proteínas y ARN son liberados. Utilizando como patrón el ARN se sintetiza una copia de ADN idéntica, mediante la transcriptasa reversa que contiene el virus (pol).Este ADN puede integrarse y permanecer en el ADN cromosómico. El proceso puede ocurrir en sentido inverso (flechas negras), en el caso de la activación de RV endógenos o exógenos durante su replicación activa, dando lugar a copias de ARN, ADN, proteínas y nuevas partículas retrovirales.
Los defectos en las actividades de los RV derivan de defectos en alguno de estos genes. Por ejemplo, defectos en el gen pol pueden afectar a la actividad integrase y hacen que un RV sea incapaz de integrarse de forma estable en el genoma tras infectar las células.
En ambos extremos del genoma de un RV existe una larga secuencia de bases que se repite en la misma orientación. Esta secuencia redundante, que contiene fundamentalmente secuencias reguladoras, se denomina LTR (Long terminal repeat). Su presencia flanqueando un gen o grupo de genes es exclusiva de los RV, ya que las LTR se forman por el modo de replicación de su genoma mediante la retrotranscripción del ARN. Esto supone para los investigadores una peculiar seña de identidad que indica el origen retroviral de un ADN desconocido.
La enorme variedad de estos agentes hace que su clasificación sea compleja, ya que atiende a diferentes propiedades de los mismos. Por ejemplo, la interacción de los RV con el huésped es muy variable y va desde la inducción rápida de tumores o SIDA a la presencia completamente benigna e incluso beneficiosa. También el ciclo replicativo y la morfología son variables que se emplean para su clasificación (tabla 1).
Tabla 1. Clasificación de los retrovirus
|
Ciclo biológico |
Patogenicidad |
Estructura viral |
|
Endógenos (HRES) |
Oncovirus:inducen |
A:estrictamente intracelular |
|
Exógenos (VIH) |
Lentivurus:patógenos |
B:extracelular con espinas |
|
- |
Espumavirus: inducen |
C:se ensamblan en la |
|
- |
- |
D: intracelular y |
HRES: retrovirus endógenos humanos VIH- I: virus de la inmunodeficiencia humana.HTLV: virus linfotrópico T humano. HSRV: retrovirus espuma humano.
![]()